上海實驗科研技術(shù)服務(wù)服務(wù)
轉(zhuǎn)錄組測序結(jié)果及TCGA數(shù)據(jù)庫分析)圖5RNA-Seq和m6A-seq聯(lián)合鑒定SOCS2是介導(dǎo)的m6A修飾的下游靶基因PLoSOne2015,在許多不同種類的RNA中,都已觀察到N6-腺苷(m6A)的甲基化,但其在microRNAs中還沒有被研究。研究者在FTO1C1,FTO2D4和FTO3C3細(xì)胞系中,通過敲除m6A甲基轉(zhuǎn)移酶FTO篩選到表達(dá)差異的microRNA,說明miRNA受m6A甲基化的調(diào)控。進(jìn)一步通過MeRIP-Seq發(fā)現(xiàn)相當(dāng)一部分的microRNA具有m6A修飾。通過motif分析,他們發(fā)現(xiàn)了區(qū)分甲基化和非甲基化microRNA的一致序列。該文章所述的表觀遺傳修飾在基因表達(dá)的轉(zhuǎn)錄調(diào)控的復(fù)雜性上增加了一個新的層次。圖FTO敲除對甲基化的miRNAs的穩(wěn)定狀態(tài)的影響。參考文獻(xiàn)Y,DominissiniD,RechaviG,HeC:Geneexpressionregulationmediatedthroughreversiblem(6)(5):(1):(12):(6):(1):(1):(uridine)(41):(6)(7540):(1):(7481):(4):(6)A-LAIC-seqrevealsthecensusandcomplexityofthem(6)(8):UTRm(6)(4):(7544):(6)(6):(5):(7667):(2):"">panstyle="color:#f5c81c;">xiainducesthebreastcancerstemcellphenotypebyHIF-dependentandALKBH5-mediatedm(6)(14):"">panstyle="color:#f5c81c;">(40):(6)(3):(1):(1):(4):(11):。純干貨Western blot (WB)條帶灰度統(tǒng)計與GraphPad作圖.上海實驗科研技術(shù)服務(wù)服務(wù)
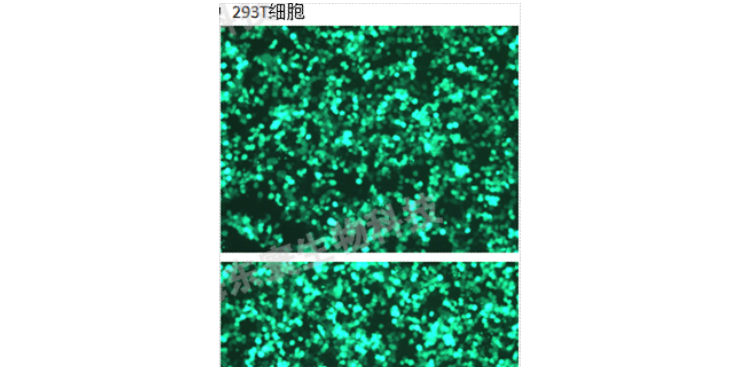
用途基因功能研究、免/殺傷/增殖等、抗體活性篩選(細(xì)胞水平的結(jié)合和阻斷)、CAR分子的殺傷活性評價。材料與儀器(以慢pMSCV載體為例)包含目的基因和eGFP-Tag的pMSCV質(zhì)粒、帶有eGFP-Tag的pMSCV空載質(zhì)粒、GAG質(zhì)粒、VSV質(zhì)粒、Puromycin、Polybrene、Lipo3000、HEK293T細(xì)胞、opti-MEM、DMEM、FBS、雙抗、μm的濾膜、熒光顯微鏡等。步驟1、基因的構(gòu)建1)根據(jù)目的基因mRNA編碼區(qū)設(shè)計引物,分別在引物兩端加入酶切位點EcoRI和BglII。2)從細(xì)胞中提取目的基因mRNA,然后逆轉(zhuǎn)錄成cDNA,然后從cDNA里面用引物把目的基因的CDS區(qū)擴(kuò)增出來。PCR擴(kuò)增出帶有酶切位點的目的基因編碼區(qū)序列,連接至pMD19-T載體后轉(zhuǎn)化至感受態(tài)DH5α,分別進(jìn)行菌落PCR鑒定和酶切鑒定。3)使用EcoRI和BglII雙酶切下目的基因序列,電泳割膠回收純化,連接到pMSCV-eGFP載體。再次轉(zhuǎn)化到感受態(tài)DH5α,菌落PCR鑒定和酶切鑒定成功后,送至公司測序、鑒定。4)鑒定成功后,將質(zhì)粒轉(zhuǎn)化至感受態(tài)DH5α中,并進(jìn)行無內(nèi)質(zhì)粒抽提。GAG質(zhì)粒和VSV質(zhì)粒同樣可以轉(zhuǎn)化至感受態(tài)DH5α中,并進(jìn)行無內(nèi)質(zhì)粒抽提。質(zhì)粒抽提后冷凍于-20℃保存。2、慢包裝1)使用DMEM完全培養(yǎng)基培養(yǎng)6cm皿HEK293T至匯合度為70~80%。上海動物科研技術(shù)服務(wù)培養(yǎng)EdU細(xì)胞增殖檢測是一種新型的細(xì)胞增殖檢測方法。

膿毒癥動物模型必須具備以下基本要素:有膿毒癥典型的高排低阻血流動力學(xué)表現(xiàn)和高代謝狀態(tài);伴發(fā)多個功能障礙;有較高的自然死亡率,根據(jù)膿毒癥的轉(zhuǎn)歸,要求動物模型的自然死亡率達(dá)到50%~70%;膿毒癥是嚴(yán)重引起機(jī)體的炎癥反應(yīng)過度造成的自身損傷,不是細(xì)菌和內(nèi)對機(jī)體的直接損傷,故出現(xiàn)功能障礙及動物死亡距膿毒癥模型制備應(yīng)有一定的時間間距。一般在制模后6~12h后發(fā)生的功能障礙或死亡屬全身炎癥反應(yīng)所致。實驗動物的選擇在制作動物模型時多選用雄性小鼠,因為雌性小鼠較雄性小鼠更能耐受膿毒癥和失血性休克,且進(jìn)入發(fā)期的雌性小鼠性水平變化很大,而雄性小鼠在膿毒癥時更易于發(fā)生免疫抑制。為什么選擇CLP模型?盲腸結(jié)扎穿孔模型(CLP)模型是接近于人類膿毒癥機(jī)制的模型,被稱為膿毒癥模型的“金標(biāo)準(zhǔn)”。CLP技術(shù)在20世紀(jì)70年代被建立。CLP模型非常適宜用于防治膿毒癥或膿毒性休克新藥的臨床前觀察造模方法CLP膿毒癥模型的建立主要分為兩個階段:盲腸遠(yuǎn)端結(jié)扎和盲腸穿刺。首先是手術(shù)引發(fā)的結(jié)扎部位組織變性壞死引起局部炎癥反應(yīng),其次是穿孔后使糞便內(nèi)容物漏入腹膜引起多菌性細(xì)菌性腹膜炎,進(jìn)而誘發(fā)全身性炎癥反應(yīng)。
動物模型在科研中有著普遍的應(yīng)用。首先,它們可以幫助科研人員深入理解的共同性,即不同物種之間存在的共有理變化過程。通過對動物模型的研究,科研人員可以更清楚地了解的發(fā)展過程和機(jī)制,為人類的檢查提供理論依據(jù)。其次,動物模型還為新研發(fā)和苗測試提供了的平臺。在研發(fā)過程中,科研人員可以通過對動物模型進(jìn)行處理,觀察其和副作用,為新的臨床試驗提供依據(jù)。而在苗測試中,動物模型則可以用來評估苗的性和安全性。此外,動物模型還為科研人員提供了研究人類的跨學(xué)科方法。例如,通過比較人類和動物模型的基因組學(xué)、蛋白質(zhì)組學(xué)等數(shù)據(jù),可以發(fā)現(xiàn)與發(fā)生相關(guān)的關(guān)鍵基因和蛋白質(zhì),從而為的和檢查提供新的思路。雖然動物模型在科研中發(fā)揮了巨大的作用,但也存在一些挑戰(zhàn)。首先,由于物種差異的存在,動物模型的表現(xiàn)與人類可能存在差異,因此需要謹(jǐn)慎使用。此外,動物模型的倫理問題也不容忽視,科研人員需要在符合倫理規(guī)定的前提下進(jìn)行相關(guān)研究。盡管存在挑戰(zhàn),動物模型的發(fā)展前景仍然值得期待。隨著科技的不斷進(jìn)步,科研人員將能夠開發(fā)出更為精確、實用的動物模型。動物疾病模型可以分為兩種類型:自然模型和人工模型。
METTL3能夠促進(jìn)肺腺細(xì)胞的生長、生存和侵襲,但還不清楚它是否作為m6A調(diào)節(jié)器或效應(yīng)器發(fā)揮作用[25]。在急性髓細(xì)胞白血病(AML)患者中,m(6)A調(diào)控基因的突變或拷貝數(shù)變化與TP53突變存在密切聯(lián)系,且m(6)A調(diào)控基因的改變與AML不良預(yù)相關(guān)[26]。此外,F(xiàn)TO在AML中高表達(dá),它通過降低mRNA轉(zhuǎn)錄本中的m(6)水平,調(diào)節(jié)ASB2和RARA等靶點的表達(dá),增強(qiáng)了白血病基因介導(dǎo)的細(xì)胞轉(zhuǎn)化和白血病形成,并抑制全反式維甲酸(ATRA)誘導(dǎo)的AML細(xì)胞分化[27]。在脂肪形成過程中,F(xiàn)TO表達(dá)與m6A水平成負(fù)相關(guān),促進(jìn)脂肪形成[3]。在膠質(zhì)細(xì)胞瘤樣細(xì)胞中,ALKBH5通過lncRNAFOXM1介導(dǎo)FOXM1基因pre-mRNA上的m6A修飾維持膠質(zhì)瘤細(xì)胞的成瘤性[28]。此外,甲基轉(zhuǎn)移酶METTL3或METTL14的敲除,能夠改變m6A的富集和ADAM19的表達(dá),極大地促進(jìn)了膠質(zhì)瘤細(xì)胞的生長、自我更新和形成[29]。圖2m6ARNA修飾和介導(dǎo)的功能[30]m6A的研究方向主要是通過研究m6A修飾相關(guān)的甲基化、去甲基化酶和識別蛋白的功能,進(jìn)而研究m6A修飾的生物學(xué)功能和作用機(jī)制:一般通過敲除m6A酶分子,研究下游功能基因分子的表達(dá)和m6A甲基化情況,通過介導(dǎo)相關(guān)基因異常(可變剪切、穩(wěn)定性、翻譯、miRNA調(diào)控)影響細(xì)胞表型和功能特征。瘤細(xì)胞的增殖活性,從而更好地評估腫、瘤的惡性程度和預(yù)后.云南動物科研技術(shù)服務(wù)外包
干貨分享 | 瓊脂糖核酸電泳,選擇紫外還是藍(lán)光?上海實驗科研技術(shù)服務(wù)服務(wù)
RNA各種可逆的化學(xué)修飾被認(rèn)為是一種新的表觀遺傳調(diào)控方式。m6A是真核生物mRNA常見的化學(xué)修飾,在調(diào)控mRNA穩(wěn)定性,剪切和翻譯方面具有重要的作用。作者使用轉(zhuǎn)錄組測序發(fā)現(xiàn)了METTL3(甲基轉(zhuǎn)移酶3),一種主要的RNAN6-腺苷-甲基轉(zhuǎn)移酶,在人肝細(xì)胞(HCC)和多種實體中高表達(dá)。在臨床上,METTL3的過度表達(dá)與肝細(xì)胞患者不良預(yù)有關(guān)。體外實驗證明敲除METTL3會抑制HCC細(xì)胞增殖,遷移及克隆形成。體內(nèi)實驗證明敲除METTL3會明顯抑制HCC體內(nèi)成瘤和肺轉(zhuǎn)移。另外,使用CRISPR/dCas9-VP64系統(tǒng),內(nèi)源性高表達(dá)METTL3會促進(jìn)HCC細(xì)胞在體外和體內(nèi)生長。通過轉(zhuǎn)錄組測序、m6A-Seq、MeRIP-PCR,作者確定了SOCS2(細(xì)胞因子信號2的抑制因子)作為METTL3介導(dǎo)的m6A修飾的下游靶基因。敲除METTL3表達(dá)會消除SOCS2mRNAm6A修飾并增強(qiáng)SOCS2mRNA表達(dá)。m6A介導(dǎo)的SOCS2mRNA降解是依賴于m6A“讀取器”蛋白YTHDF2。總之,METTL3在HCC大部分高表達(dá)中,并通過m6A-YTHDF2依賴機(jī)制抑制SOCS2表達(dá)從而促進(jìn)HCC進(jìn)展。因此,作者發(fā)現(xiàn)了在肝發(fā)生過程中表觀遺傳改變的一種新機(jī)制。圖4RNA甲基化轉(zhuǎn)移酶METLLT3在肝組織中高表達(dá)。上海實驗科研技術(shù)服務(wù)服務(wù)
- 云南裸鼠原代細(xì)胞實驗 2025-10-13
- 湖北哪里有原代細(xì)胞分離 2025-10-13
- 西藏裸鼠動物模型培養(yǎng) 2025-10-13
- 上海豚鼠動物模型手術(shù) 2025-10-13
- 江蘇組織原代細(xì)胞技術(shù) 2025-10-12
- 湖北C57原代細(xì)胞構(gòu)建 2025-10-12
- 湖北組織原代細(xì)胞技術(shù) 2025-10-12
- 皮下成瘤動物模型實驗室 2025-10-12
- 云南小鼠原代細(xì)胞服務(wù) 2025-10-12
- 哪里有原代細(xì)胞 2025-10-12
- 深圳NIKE驗廠輔導(dǎo) 2025-10-13
- 昆山參考企業(yè)形象策劃聯(lián)系人 2025-10-13
- 四川品牌電商配送服務(wù)24小時服務(wù) 2025-10-13
- 貴陽好玩的露營選擇 2025-10-13
- 杭州高層外墻清洗哪家公司性價比高 2025-10-13
- 延慶區(qū)專業(yè)的支原體檢測平臺 2025-10-13
- 山西哪些進(jìn)出口資質(zhì)辦理 2025-10-13
- 閔行區(qū)汽車零配件銷售工廠 2025-10-13
- 沭陽創(chuàng)新文化活動策劃五星服務(wù) 2025-10-13
- 成都信息化系統(tǒng)性能檢測 2025-10-13